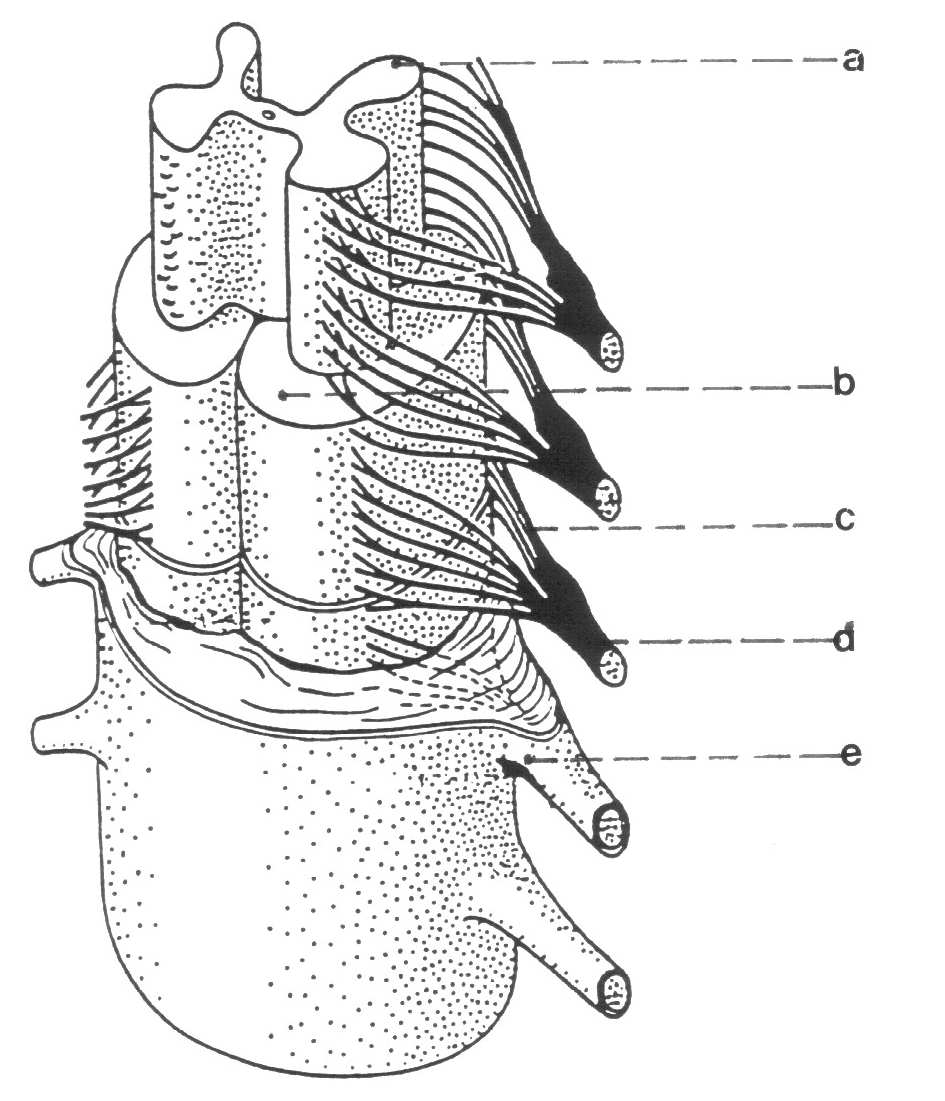
Obr. 3. 3.
Prostorová rekonstrukce míchy
Prostorová rekonstrukce míchy
- šedá míšní hmota
- bílá hmota
- zadní míšní koren
- míšní nerv
- míšní obaly
Reflex je funkční jednotkou nervové soustavy. Reflex je odpovědí organizmu na podnět, podráždění, změnu zevního nebo vnitřního prostředí. Tato odpověď se uskutečňuje na určité anatomické struktuře, která je daná strukturou reflexního oblouku.
Reflexní oblouk se skládá z receptoru, aferentní dráhy (aferentního, přívodného neuronu), eferentní dráhy (eferentního neuronu) a efektoru (svalu). Nás budou zajímat reflexní oblouky uplatňující se při řízení pohybu.
Je-li mezi aferentním neuronem a motoneuronem (eferentní neuron) pouze jedna synapse (oblouk tvoří jen dva neurony), jde o monosynaptický reflexní oblouk, jsou-li mezi aferentní a eferentní neurony vsunuty další neurony (interneurony), jde o polysynaptický reflexní oblouk. (Počet vsunutých interneuronů může být velmi různý - od jednoho do několika tisíc.)
Mícha je nejnižším reflexním ústředím centrálního nervového systému.
Míšní reflexy dělíme podle receptorů (proprio, exteroceptorů), jejichž podrážděním můžeme příslušnou odpověď vybavit na :Receptory proprioceptivních reflexů jsou svalová vřeténka a šlachová tělíska.
Reflexní oblouk proprioceptivního reflexu tvoří svalové vřeténko nebo šlachové tělísko => aferentní nervové vlákno míšního nervu (buňka ve spinálním gangliu) => alfa motoneuron (buňka v předním míšním rohu) => efektor (kosterní sval).
Jak vyplývá z popisu reflexního oblouku, jde o velmi jednoduchý, někdy i monosynaptický oblouk, který je proto velmi rychlý a má mimořádně krátkou reflexní dobu odpovědi – 10 ms.
Svalové napětí (svalový tonus) je výchozím předpokladem pro provedení jakéhokoliv pohybu a pro udržení vzpřímené polohy těla.
Název proprioreceptivní (tj. vlastní), vyjadřuje skutečnost, že podráždění přicházející z určitého svalu je převedeno na motoneuron téhož svalu. Reflexní oblouk tedy začíná a končí v tomtéž svalu.
Informace přicházející ze svalových receptorů nekončí jen u motoneuronů předních míšních rohů (monosynaptické zapojení), ale jsou předávány i do retikulární formace (RF) mozkového kmene, do mozečku, thalamu a do mozkové kůry.
Svalová vřeténka jsou několik milimetrů velké útvary, uložené při přechodu šlachy do svalu. Jemným vazivovým pouzdrem bývají oddělena od okolního vaziva svalu.
Vřeténko se skládá z 6–8 tenkých svalových vláken, která jsou 2–10 mm dlouhá. Svalovým vláknům vřeténka se říká intrafuzální. Vlastní vlákna kosterního svalu jsou tedy vlákny extrafuzálními. Oba typy vláken jsou ale uspořádány paralelně! Na zevním obvodu intrafuzálních vláken jsou typické motorické ploténky gama motoneuronů. Intrafuzální vlákna tedy mají samostatnou motorickou inervaci a ve vřeténcích začínají i dva typy aferentních, senzitivních nervových vláken. Tato vlákna spirálovitě obtáčejí intrafuzální vlákna vřeténka jdou jako aferentní vlákna míšního nervu a zadními míšními kořeny vstupují do míchy. Těla (perikarya) těchto neuronů jsou uloženy ve spinálních gangliích.
Vřeténka jsou drážděna při protažení svalu. Tím, že intrafuzální vlákna vřeténka probíhají paralelně s ostatními svalovými vlákny, jsou natahována současně s prodloužením svalu a proud vzruchů zpětnovazebně dráždí alfa motoneurony vyvolávající svalovou kontrakci. Vřeténka jsou drážděna i váhou končetin a tahem antagonních svalů. Zajišťují kontakt kloubních ploch a postavení kloubu, ale svalovou kontrakci přímo neregistrují!
Vřeténko je jakýsi komparátor, který srovnává napětí intrafuzálních vláken a vláken svalu. Gama inervace intrafuzálních vláken navozuje jejich kontrakci a určuje tak jejich předpětí – nastavení vřeténka na určité napětí.
Svalová vřeténka jsou svým zapojením autoregulační systém, jehož nastavení je významně ovlivňováno (přes interneurony) retikulární formací mozkového kmene. Celému systému této zpětnovazebné inervace říkáme gama smyčka (klička).
Část aferentních vláken přicházejících ze svalových vřetének, je zapojena i na alfa motoneurony antagonních svalů. Pomocí tohoto zapojení dochází při kontrakci agonistů a synergistů ke ztlumení napětí antagonistů. Bez jejich „vypojení“ by totiž nebylo možné uskutečnit žádný koordinovaný pohyb. Vypojení antagonistů pomocí této, tzv. reciproční inervace, zajišťují především míšní interneurony. Utlumení antagonistů pomocí reciproční inervace není ale nikdy úplné. „Zbytková“, neutlumená aktivita je pro funkci pohybového aparátu dokonce výhodná – chrání kloubní pouzdra a vazy před prudkými „bičovými“ pohyby a dovoluje rozsah pohybu plynule dávkovat.
Šlachová tělíska (Golgiho tělíska) jsou drobné receptory, uložené v průběhu šlach, vazů a kloubních pouzder. Tělísko tvoří několik svazků kolagenních vláken, která opřádají bohatě rozvětvená aferentní nervová vlákna. Tělísko je obaleno jemným vazivovým pouzdrem.
Aferentní nervová vlákna šlachových tělísek mají buňky ve spinálních gangliích. Axony těchto buněk jdou do míchy k interneuronům, prostřednictvím kterých tlumí aktivitu alfa motoneuronů inervujících kosterní svaly.
Šlachové tělísko chrání kosterní sval před přetažením. Tělísko je tedy aktivováno při protažení svalu (tahem za šlachu) i při svalové kontrakci (opět tahem za šlachu), kterou svalové vřeténko nezaznamenává.
Tělíska jsou na svalovou kontrakci citlivější než na pasivní protažení šlachy. Souhrou činnosti vřetének a tělísek je tak zajištěna dokonalá informace centrálního nervového systému o napětí, stupni kontrakce i zatížení všech svalů.
Uvědomování si svalového napětí, polohy končetin a trupu, změny polohy a rychlostí této změny nám umožňuje tzv. hluboký svalový smysl. Je to složitý a nepřesně definovatelný vjem, který zajišťuje souhra svalových vřetének, šlachových tělísek a receptorů kloubních pouzder a vazů atd. Tohoto komplexního vjemu se účastní i zrak, sluch, orgány rovnováhy. Klíčový pro tento vjem je zřejmě zpětnovazebný mechanizmus řízení svalové kontrakce pomocí gama smyčky.
Receptory exteroreceptivních reflexů jsou receptory pro bolest a dotyk, které jsou uloženy v kůži. Podle odpovědi, kterou na podráždění dostaneme, rozlišujeme extenzorové a flexorové reflexy.
